August Krogh
Nobel Prize lecture
Nobel Lecture, December 11, 1920
A Contribution to the Physiology of the Capillaries
At the beginning of the 17th century, Harvey succeeded in demonstrating that the blood within an organism was in constant circulation, passing out from the heart, through the arteries, to all the various organs, and returning through the veins. About 50 years later, Malpighi, (and, at about the same time, Leeuwenhoek), discovered that the connection between the smallest branches of the arteries and the veins was not by random cavities between the organs, but by a network of extremely fine vessels – the capillaries, visible only under the microscope. These have therefore been known for about 250 years, and it had been obvious, almost from the first, that they were, in one sense, the most important element in the whole circulatory system. All exchange between the blood and the organs, of nutrient materials and waste products, of oxygen and carbon dioxide, takes place through their walls. In them the blood fulfils its real function. One could say that the whole circulatory machine exists for their sake. It is all the more remarkable that their physiological behaviour has not been investigated to any extent, and even now is comparatively little known, so little known in fact that it has been possible for me to make original contributions by nothing more than patient observation under the microscope and simple probing with fine needles.
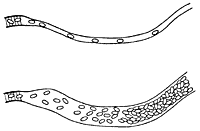
I believe that it is easiest for me to present the results of my investigations in this field by setting forth, and dealing with, the problems in the order in which they occurred to me. I therefore begin by discussing the provision of oxygen for the muscles. In a muscle the capillaries run along the length of the muscle fibres, as shown in Fig. 1, and the oxygen in the blood has to diffuse through the wall of the capillary into the muscle substance.
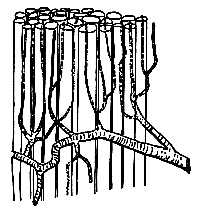
Through comparing various researches, some by Verzàr, some by Lindhard and myself, into the utilization of oxygen by muscles and its dependence, partly on the oxygen pressure in the blood, partly on muscular activity, I came, in 1915, to the conclusion that, either there must be very serious errors in the researches concerned, or the circulation in the capillaries must otherwise occur in an entirely different way from that which one normally visualized. The conventional picture is that shown in Fig. 1. All the capillaries are open, and of about the same width, so that the blood passes through all of them at about the same rate. This rate was thought to be regulated by the greater or lesser contraction of the small arteries, which convey blood to the capillaries. When the small arteries constricted, and offered high resistance, the blood came to flow slowly through all capillaries. When they dilated, the blood flow increased very strongly, as was known to occur during muscular activity.
The conclusion to which I was forced, in order to reconcile the results of earlier researches, was this, that in a resting muscle the blood could not flow through all of the capillaries, but only through a small percentage of them, which, further, must be more or less regularly distributed throughout the muscle, as is shown in Fig. 2, which is a wholly schematic representation of capillaries in a cross-section of muscle. This conclusion was rather daring at that stage, because it necessarily implied that the capillaries were not, as had hitherto been supposed by nearly everyone, tubes which quite passively permitted the passage of blood, and whose bore was determined by their internal pressure, but must have independent powers of contraction. It was therefore essential to test this assumption experimentally before one could give it any credence.
 |
| Fig. 2. Cross-section of muscle showing open O and closed • capillaries. |
By direct observation, under the microscope, of living muscle, some from the frog, some from small mammals, it was comparatively easy to determine that the number of visible capillaries, which is the same as the number of capillaries through which blood flows, was rather small in resting muscle, and only increases very appreciably when the muscle has been active for some seconds. However, the other important question, as to whether the open capillaries were approximately regularly distributed, or whether perhaps they became empty of blood by groups, when the arterial branch which supplied the group closed, this was not so easy to answer.
Here I had to begin by counting the capillaries. When one had filled the vascular system by injecting a strongly coloured medium which later hardens, one can make preparations which show the filled vessels with very great clarity. Fig. 3 shows in this way an extremely coarse capillary network from the stomach of the frog. Fig. 4 is a cross-section, approximately 0.5 mm in diameter, of muscle from the horse, in which the individual capillaries show themselves as small black dots between the muscle fibres, and from such a preparation one can count directly how many capillaries are found in a cross-section of a given size, say, one square mm. The counts I have carried out now show that the numbers vary appreciably in different animals, being comparatively low in the cold-blooded, high in the warm-blooded, particularly in small animals, where in, for example, a guinea-pig, one finds approximately 3,000 to the sq. mm, that is to say, in a cross-section of an area little larger than that of a pin’s head. Because the length of each capillary is, on average, only 0.5 mm, the total number of capillaries in a large organism, such as the human, is incredibly large, and one can calculate, for example, that, if all the capillaries in a grown man formed a continuous tube, it could reach at least twice round the earth.

 |
| Fig. 5. Longitudinal section of the sartorius muscle of the frog. (Vital injection, x 100.) |
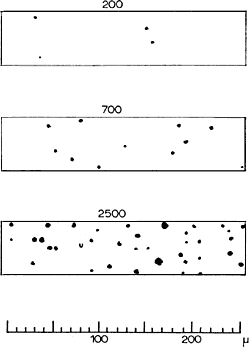
In order to ascertain which of these many capillaries are open, and which are closed, at any one moment in the living organism, I dispersed a suspension of microscopic black particles (Indian ink was found to be the most suitable), in the blood of a living animal, by intravenous transfusion. When the animal is killed shortly afterwards, one finds Indian ink in those capillaries which were open at that particular moment, while the others will not be visible. Fig. 5 shows a muscle preparation from a frog, which had been subjected to muscular work immediately before transfusion with Indian ink, and in which, therefore, numerous capillaries were open. One can clearly see, in several places, the uncoloured corpuscles, outlined by the black-coloured blood plasma. The difference between active and resting muscle is brought out particularly clearly in such preparations, and it was found that, whether the open capillaries are few or numerous, their distribution is always fairly regular. Fig.6 shows an optical cross-section of a small area, ca. 1/80 mm2, of three different muscles from the same guinea-pig. In the top specimen there are only 3 capillaries visible (a total of 200 per sq. mm). In the bottom specimen, which is from a respiration muscle which had been subjected to work right up to the moment the animal was killed, one finds 2,500 open capillaries per sq. mm, or, in other words, nearly all the existing capillaries were in use. The illustration further shows that there is a great difference in the bore of the capillaries which are open. In resting muscle, all are very narrow, but in active muscle a number are rather wide. Comparison with the scale, which is marked in thousandths of a mm, shows that the diameter can decrease down to 2-3 thousandths of a mm. It is remarkable that blood corpuscles, which are round discs of 7 ![]() diameter, and 2
diameter, and 2 ![]() thickness, can pass through such narrow channels, but Fig. 7 shows this can occur, since they either roll up, or simply elongate into long sausage-shaped bodies.
thickness, can pass through such narrow channels, but Fig. 7 shows this can occur, since they either roll up, or simply elongate into long sausage-shaped bodies.
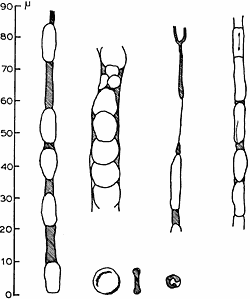 |
| Fig. 7. Various capillaries from guinea-pig muscle. (Vita injection.) |
It clearly follows from the researches, of which I have now given a short account, that capillaries, at any rate in muscle, must obviously have powers of independent contraction, that, in other words, there must be a special capillary-motor system by which the bore of the capillaries can be regulated, but stating that fact raises a whole series of new questions: is variation in diameter of capillaries independent of the arteries, or does it follow from them? In what way can the capillaries be excited – chemical, electrical or mechanical? Are they under nervous control, and, if so, by which nerve? Can contractile histological elements be demonstrated in their walls? And so on. I have been engaged on these and other related questions for the past few years; research is in full progress at this moment, and there still seems so much to be done that I am tempted to use the metaphor from one of the works of my great fellow countryman, Nicolaus Steno – that our misgivings are like the heads of the Hydra of Lerna – every time one is cut off, several new ones arise. When one is in the middle of such work, where the working hypotheses are subject to constant change, it is extremely difficult to give a coherent report, and I must content myself with some examples of what has been achieved, and hint at conclusions which I hope to reach.
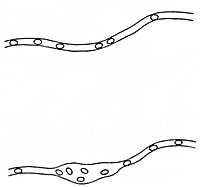
First, I have attempted to solve the question as to whether capillary changes in the tongue of the frog are independent of the arteries. When the tongue of a frog is spread across a glass slide, it is a particularly suitable specimen for investigation. It is translucent and, on one side, a smooth mucous membrane is found, with a widely distributed capillary network, in which the individual capillaries are readily responsive to excitation, and where both the small arteries and veins can be observed directly. So long as the tongue is not excited, the great majority of these capillaries are closed, and the tongue is extremely pale and bloodless. It is well known that mechanical irritation of human skin by, for example, a scratch from a needle, produces a red streak, because the vessels fill with blood. A corresponding reaction can be produced very easily on the tongue of the frog, and under the microscope one can show that this depends, in the first instance, on capillary dilation, and one can bring about dilation of a single capillary, or part of a capillary. Fig. 8 shows such a capillary, part in uninfluenced condition, part after it has been lightly scratched in the middle. Fig.9 shows how it is further possible to find a completely closed capillary by careful scratching along a tiny venous branch (v). Blood passes out from the vein, and fills a part of the capillary, but does not flow through. By further scratching, one can obtain the capillary filled bit by bit, until it makes connection with an open capillary or artery, when flow suddenly commences. This experiment is important because it shows that the very low venous pressure is sufficient to fill capillaries whose walls are flaccid, while the high arterial pressure cannot force entry into a contracted capillary.
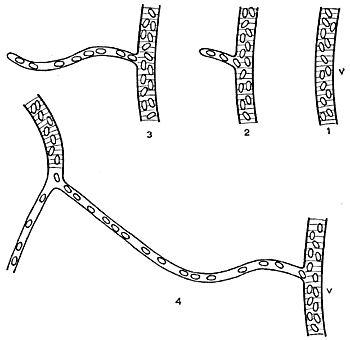 |
| Fig. 9. Effect of repeated mechanical irritation on the tongue of the frog (v = tiny venous branch). |
This behaviour is of great theoretical and practical importance. One can conclude from this and from many other analogous facts that, when appreciable capillary dilation occurs, it cannot be because of a simple rise in arterial blood pressure but must depend on change in condition of the capillary walls – a relaxation of their contractile elements. Widening of the arteries, on its own, only leads to a higher pressure in the capillaries, and a more rapid flow of blood through them. The greater or lesser red colouration of an organ depends, in the first instance, on the blood content of the capillaries, and one is therefore justified in concluding that, in all such cases, where there is marked redness, we are dealing with widened capillaries. However, only closer examination can give information as to whether, at the same time, the arteries are widened, and capillary pressure is also high. The capillaries in human skin become, for example, widened with strong heat and by excitation with strong light, and with different emotions, while cooling to a certain level and psychological depression results in contracted capillaries. The detailed mechanism of these reactions has not yet been studied. By various chemical agents, the capillaries can be made to dilate, and urethane, for example, in fairly strong solution, causes very great dilation of the capillaries of the frog’s tongue, while it has no effect on the arteries. With urethane, one can produce the situation depicted in Fig. 10, where blood is pumped out from a very narrow artery into a capillary, which widens more and more, and retains all the blood corpuscles it receives. The capillary walls widen so forcefully that they become permeable to blood plasma, so that the capillary is at last filled by a solid mass of corpuscles. This observation is not irrelevant to the problem of oedema, and is at the moment being more closely investigated by my colleague, Dr. Harrop, along the lines of a determination of the size of openings which appear in the walls of the capillaries. Suitable substances are introduced into the blood through a vein. A group of capillaries in the tongue of the frog can be caused to dilate themselves, and observations made on the substance passing out through the capillary walls. If the colloidal dye “vital red” was administered, followed by application of urethane, a fine red stripe appeared alongside the widened capillary. If Indian ink, whose particles are on the borderline of the microscopically visible, was administered, the widened capillaries showed themselves closed towards that substance. We have similarly ascertained that molecules of starch can pass through.
A reaction corresponding to urethane dilation in the tongue of a frog is known from human skin, when it becomes reddish-blue under great cooling. We then also have strongly dilated capillaries and constricted arteries, so that the blood flow becomes very slow, and an essential part of the blood oxygen is used up during its passage, resulting in the blue colouration. This reaction is in line with many other vasomotor reactions in human skin, closely studied by Ebbecke, who described them concurrently with the commencement of my capillary researches, and derived a series of important conclusions. I am particularly anxious to emphasize Ebbecke’s valuable work because, in my first report, I was not aware of its existence.
A very large number of substances are found which produce capillary dilation and, by stronger action, oedema. Thus it is only necessary for me to consider the substances producing inflammation. I must draw attention, in this connection, to histamine, which Dale, in a very beautiful investigation, has shown to produce capillary dilation in various mammals. In the case of the frog’s capillaries, histamine is virtually without action, and this fact illustrates in a striking way the observation that all capillaries do not at all show the same reactions. Even in the same animal there is a very great difference, which has naturally complicated the study, and added to its interest.
I have closely investigated conditions in the skin and in the web of the frog. Here the capillary system is entirely different from that of the tongue. The capillary network is very dense and most capillaries are always open, though narrow. They react only weakly to chemical reagents which produce strong dilation in the tongue, and while very slight mechanical irritation can cause some dilation, as a rule they contract when the excitation is stronger. This is of interest because so far there has been no success in finding any stimulus which could produce contraction in the capillaries of the tongue.
I describe as the capillaries’ tonus the fact that they have the power of contracting to a greater or lesser extent, and of maintaining themselves contracted. In the frog’s tongue, capillary tonus is very strongly developed, in that by far the majority are completely contracted. In the web, it is much weaker, in that the capillaries are only exceptionally completely closed. Most muscle capillaries take up an intermediate position. It can very easily be shown that blood flow is a condition for the maintenance of tonus, but that dependence on the blood is very varied. For example, if one stops the blood supply to the web for 10-20 minutes, all the capillaries dilate very strongly as soon as the blood is allowed to enter. They thus relax very rapidly when there is not a steady stream of fresh blood through them. If one performs the same experiment with the tongue, many hours must elapse before a similar reaction occurs, and even the slightest blood supply is adequate to maintain tonus. When the blood flow is restored following obstruction, marked hyperaemia results. This fact has long been known, and utilized in therapy. One has assumed that the alteration in the exchange of substances, which leads to oxygen deficiency and the production of acidic substances, was the causative factor. This explanation, at least so far as concerns the organs which I have investigated, is quite incorrect. They receive all the oxygen they require from the atmosphere, and, apart from this, we have found that much higher degrees of acidity than ever occur in the tissues are required to cause even the tongue’s sensitive capillaries to dilate. The explanation must be sought in an entirely different direction. The blood must contain a substance which acts on the contractile elements in the capillary walls, and stimulates them to contract. To investigate, and if possible isolate this substance, is one of the problems which will be taken up in the near future.
The presence of such a substance in the blood implies an extremely complete, regulatory mechanism, by which the blood is quite regularly distributed in the most economic way. If a capillary has been restricted of blood for a certain time, its tone will diminish, and blood will flow through until it can, once again, close itself. The open capillaries in the tongue or in muscle will thus continuously shift their place. Tissue elements which are at a given moment situated far from open capillaries and are therefore poorly supplied will receive blood passing close by a little while later. No part of the tissues will be deprived, and the blood is utilized in the most economic way.
Time does not permit me to go into the conditions of the innervation of the capillaries, which shows many interesting features, but which, as yet, lacks much towards a complete elucidation. I must also content myself with the utmost brevity concerning the demonstration of the histology of the contractile elements. Dr. Vimtrup, among my collaborators, is working on this question, and has succeeded in observing, surrounding the capillaries, the branched cells which were described by Rouget as long ago as the 70’s. However, no definitive results have yet been achieved. I think I have said enough to make it clear to you that the researches which I have explained deal with significant problems, and that it will need a great deal of work, difficult in many respects, to bring them to a satisfactory solution. The tremendous honour of the Nobel Prize is of the strongest incentive to me in my work, while the amount of the Prize will greatly simplify my task, and provide me with much valuable help in my work. I ask you to be assured that I will do all I possibly can to show myself worthy of the trust which the Caroline Institute has shown in me.
[The lecture was illustrated by a film projection.]
The Nobel Foundation's copyright has expired.Nobel Prizes and laureates
Six prizes were awarded for achievements that have conferred the greatest benefit to humankind. The 14 laureates' work and discoveries range from quantum tunnelling to promoting democratic rights.
See them all presented here.

