Thomas H. Morgan
Article
Thomas Hunt Morgan and his legacy
by Edward B. Lewis
1995 Nobel Laureate in Physiology or Medicine
This article was published on 20 April 1998.
Thomas Hunt Morgan was awarded the Nobel Prize in Physiology or Medicine in 1933. The work for which the prize was awarded was completed over a 17-year period at Columbia University, commencing in 1910 with his discovery of the white-eyed mutation in the fruit fly, Drosophila.
Morgan received his Ph. D. degree in 1890 at Johns Hopkins University. He then went to Europe and is said to have been much influenced by a stay at the Naples Marine Laboratory and contact there with A. Dohrn and H. Driesch. He learned the importance of pursuing an experimental, as opposed to descriptive, approach to studying biology and in particular embryology, which was his main interest early in his career. A useful account of Morgan’s life and works has been given by G. Allen (ref. 1).

Thomas Hunt Morgan with fly drawings.
Courtesy of the Caltech Archives. © California Institute of Technology. All rights reserved. Commercial use or modification of this material is prohibited.
In 1928 he moved with several of his group to Pasadena, where he joined the faculty of the California Institute of Technology (or Caltech) and became the first chairman of its Biology Division. What factors were responsible for the successes that Morgan and his students achieved at Columbia University and how did these factors carry over to the Caltech era first under Morgan’s, and later G.W. Beadle’s leadership? It is convenient to consider three time periods:
Morgan and the Columbia Period (1910 to 1928)
Morgan attracted extremely gifted students, in particular, A.H. Sturtevant, C.B. Bridges, and H.J. Muller (Nobel Laureate, 1946). They were to discover a host of new laws of genetics, while working in the “Fly Room,” in the Zoology Department at Columbia.
 |
 |
 |
| A.H. Sturtevant | C.B. Bridges | H.J. Muller |
|
Courtesy of the Caltech Archives
Copyright © California Institute of Technology. All rights reserved. Commercial use or modification of this material is prohibited. |
||
Throughout their careers Morgan and these students worked at the bench. The investigator must be on top of the research if he or she is to recognize unexpected findings when they occur. Sturtevant has stated that Morgan would often comment about experiments that led to quite unexpected results: “they [the flies] will fool you every time.”
Morgan attracted funding for his research from the Carnegie Institution of Washington. That organization recognized the basic research character of Morgan’s work and supported research staff members in Morgan’s group, such as C.B. Bridges and Morgan’s artist, Edith Wallace, who was also curator of stocks. The Carnegie grants required nothing more than an annual report from the investigators. Federal support had not yet started and although universities were able to finance costs associated with teaching they were usually unable to support basic research.

C. Bridges, P. Reed, T.H. Morgan, A.H. Sturtevant, E.M. Wallace.
Courtesy of the Caltech Archives. © California Institute of Technology. All rights reserved. Commercial use or modification of this material is prohibited.
During the Columbia period Morgan was clearly in his prime. His style of doing science must have been of paramount importance. He was not afraid to challenge existing dogma. He had become dissatisfied, even skeptical, of the formalistic treatment that genetics had taken in the period between the rediscovery of Mendelism in 1901 and 1909. He ridiculed explanations of breeding results that postulated more and more hereditary factors without any way of determining what those factors were. He wanted to know what the physical basis of such factors might be. At that time it was generally assumed that chromosomes could not be the carriers of the genetic information. He wanted a suitable animal and chose Drosophila, because of its short life cycle, ease of culturing and high fecundity. Also, large numbers of flies could be reared inexpensively — an important factor during this period when there were very few funds available to support basic research. Morgan was very thrifty when it came to purchasing laboratory equipment and supplies — but, according to Sturtevant, generous in providing financial help to his students. At the start of the work hand lenses were used. Only later did Bridges introduce stereoscopic microscopes. Bridges also devised a standard agar-based culture medium. Prior to that, flies were simply reared on bananas. In addition, Bridges built the basic collection of mutant stocks, mapped virtually all of the genes and later, at Caltech, drew the definitive maps of the salivary gland chromosomes. His enormous research output may in part be attributed to his being a staff member of the Carnegie Foundation with consequent freedom from teaching and other academic obligations.
Morgan’s first attempts to find tractable mutations to study were quite disappointing. Fortunately, he persevered and found the white-eyed fly1. This led to his discovery of sex-linked inheritance and soon with the discovery of a second sex-linked mutant, rudimentary, he discovered crossing over.

H. Sturtevant in the Drosophila stock room of the Kerckhoff Laboratories.
Courtesy of the Caltech Archives. © California Institute of Technology. All rights reserved. Commercial use or modification of this material is prohibited.
Sturtevant (ref. 2) has described how chromosomes finally came to be identified as the carriers of the hereditary material. In a conversation with Morgan in 1911 about the spatial relations of genes in the nucleus, Sturtevant, who was still an undergraduate, realized that the sex-linked factors might be arranged in a linear order. He writes that he went home and spent the night constructing a genetic map based on five sex-linked mutations that by then had been discovered. In 1912 Bridges and Sturtevant identified and mapped two groups of autosomal (not sex-linked) factors and a third such group was identified by Muller in 1914. The four linkage groups correlated nicely with the four pairs of chromosomes that Drosophila was known to possess. Proof that this correlation was not accidental came when Bridges used the results of irregular segregation of the sex chromosomes (or non-disjunction) to provide an elegant proof that the chromosomes are indeed the bearers of the hereditary factors or genes as they are now known. Bridges published this proof in 1916 in the first paper of volume I of the journal Genetics.
Sturtevant often commented on Morgan’s remarkable intuitive powers. Thus, Sturtevant describes how after explaining some puzzling results to Morgan, Morgan replied that it sounded like an inversion2. Sturtevant went on to provide critical evidence, purely from breeding results, that inversions do occur; it was only later that inversions were observed cytologically.
It seems clear that Morgan was not only a stimulating person but one who recognized good students, gave them freedom and space to work, and inspired them to make the leaps of imagination that are so important in advancing science.
Morgan and the Caltech Period (1928 to 1942)

Robert A. Millikan with cosmic ray equipment.
Courtesy of the Caltech Archives. © California Institute of Technology. All rights reserved. Commercial use or modification of this material is prohibited.
Morgan was invited by the astronomer, G.E. Hale, to chair a Biology Division at the California Institute of Technology (Caltech). Hale had conceived the idea of creating Caltech some years earlier and had already recruited R.A. Millikan (Nobel Laureate in Physics, 1923) and A.A. Noyes to head the Physics and Chemistry Divisions, respectively. According to Sturtevant, Morgan told his group at Columbia of Hale’s invitation and of how it was not possible to say no to Hale. Morgan accepted and came to Caltech in 1928. He brought with him Sturtevant, who came as a full professor, Bridges, and T. Dobzhansky, who later became a full professor. In addition to Sturtevant and Dobzhansky, the genetics faculty consisted of E.G. Anderson and S. Emerson. J. Schultz, who like Bridges was a staff member of the Carnegie Institution of Washington, participated in the teaching of an advanced laboratory course in genetics.
During this second period, many geneticists visited the Biology Division for varying periods of time. Those from foreign countries included D. Catcheside, B. Ephrussi, K. Mather, and J. Monod (1965 Nobel Laureate). Visiting professors included Muller and L.J. Stadler. B. McClintock (1983 Nobel Laureate) came as a National Research Fellow in the early 1930s.
Morgan was well known outside of the scientific community and attracted interesting people. Professor Norman Horowitz, who was a graduate student in the Biology Division during this period, tells me that he remembers Morgan giving a tour of the Biology Division to the well-known author, H.G. Wells.

Back (left to right): Wildman, Beadle, Lewis, Wiersma; standing: Keighley, Sturtevant, Went, Haagen-Smit, Mitchell, Van Harreveld, Alles, Anderson; seated (back row): Borsook, Emerson; (front row): Dubnoff, Bonner, Tyler, Horowitz.
Courtesy of the Caltech Archives. © California Institute of Technology. All rights reserved. Commercial use or modification of this material is prohibited.
J.R. Goodstein (ref. 3) has described how the Rockefeller Foundation and private donors provided financial support to the Biology and other Divisions during this period. Such assistance was essential at that time, since Caltech is a private institution and received no support from the state or the federal government.

Edward B. Lewis with Drosophila.
Courtesy of the Caltech Archives. © California Institute of Technology. All rights reserved. Commercial use or modification of this material is prohibited.
In the latter half of this period, Morgan returned to his interest in marine organisms and did not follow the newer developments in genetics. Instead it was largely Sturtevant who carried on the Morgan legacy as far as genetics was concerned. Sturtevant also allowed his graduate students considerable freedom to choose their thesis projects and to consult with him on those projects or indeed on any matter. I was fortunate to have been one such student, commencing in 1939. Sturtevant’s door was always open to students and faculty. I well remember Morgan coming to Sturtevant’s office to discuss matters affecting the Division.
Sturtevant told us that the award of the Nobel Prize to Morgan in 1933 was an important factor in elevating the prestige and status of the Biology Division at the Institute. At the time, the only other Nobel Laureate at Caltech was Millikan. From 1942 to 1946, the Division was managed by a committee chaired by Sturtevant.
Beadle and the Caltech Period (1946 to 1961)

Beadle and Pauling with molecular model.
Courtesy of the Caltech Archives. © California Institute of Technology. All rights reserved. Commercial use or modification of this material is prohibited.
In 1946, Sturtevant and Linus Pauling (who was awarded Nobel Prizes in Chemistry, 1954, and Peace, 1962) persuaded Beadle, who was then Professor of Biology at Stanford University, to become chairman of the Biology Division. Beadle carried on the Morgan tradition of strongly supporting basic research and maintaining a stimulating intellectual atmosphere. During the early 1930s Beadle had been a National Research Fellow in the Division. He had collaborated with Sturtevant on a monumental study of inversions and together they wrote a textbook of genetics. He had collaborated also during that time with Sterling Emerson, and with E.G. Anderson. Beadle was clearly a part of the Morgan legacy.

Beadle in lab coat.
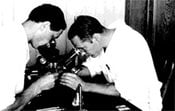
George Beadle and B. Ephrussi using microscopes.
Both photos courtesy of the Caltech Archives. © California Institute of Technology. All rights reserved. Commercial use or modification of this material is prohibited.
Beadle received the Nobel Prize in Physiology or Medicine in 1958 for work carried out at Stanford University on the biochemical genetics of the bread mold, Neurospora. In his biographical memoir on Beadle, Horowitz (ref. 4) describes how, while postdoctoral fellows in the Biology Division, Beadle and Ephrussi decided to pursue an early discovery by Sturtevant; namely, that a diffusible substance must be involved in the synthesis of the brown eye pigment of Drosophila. Sturtevant had shown that the vermilion eye color mutation is non-autonomously expressed in flies that are mosaic for the vermilion mutation and its wild-type allele. Beadle and Ephrussi designed at Caltech a set of experiments, involving transplantation of larval imaginal eye discs, to study the vermilion-plus hormone, as they called the diffusible substance. They carried out these experiments in Paris in Ephrussi’s laboratory. They were able to show that another eye color gene, cinnabar, lacks a cinnabar-plus substance and that the wild-type vermilion and cinnabar genes control sequential steps in a biochemical pathway leading to the brown eye pigment. Beadle correctly realized that the fungus Neurospora would provide better genetic material for exploring such pathways. Beadle and E. Tatum (co-winner with Beadle of the Nobel Prize) and colleagues at Stanford were then successful in dissecting the biochemical pathways that are involved in the synthesis of vitamins and many amino acids in that organism. The Neurospora findings opened a new era, now known as molecular genetics.
During Beadle’s tenure as chairman, N.H. Horowitz, H.K. Mitchell, R.D. Owen, and R.S. Edgar were added to the faculty in genetics. [I had come as an instructor in 1946 before Beadle had arrived]. Horowitz and Mitchell had been associated with Beadle at Stanford and played major roles in developing the one-gene one-enzyme hypothesis that led to the award of the Nobel Prize to Beadle and Tatum.
Beadle was responsible for persuading Delbrück to return to Caltech as a full professor. Delbrück had not been offered an appointment at Caltech after his tenure in the Division in the 1930s as a post-doctoral fellow and had taken a faculty position at Vanderbilt University. Other appointments during Beadle’s chairmanship that added strength in animal virology were R. Dulbecco (1975 Nobel Laureate), and M. Vogt. Howard Temin was one of Dulbecco’s graduate students and later a cowinner with Dulbecco of the Nobel Prize in 1975. R. Sperry (1981 Nobel Laureate) joined the faculty as a full professor in 1954 and continued his work on split brains that he had begun at the University of Chicago.

R. Dulbecco, G. Beadle, M. Delbrück and H. Temin.
Courtesy of the Caltech Archives. © California Institute of Technology. All rights reserved. Commercial use or modification of this material is prohibited.
Basic research gradually became well supported financially by Federal Agencies commencing with the Office of Naval Research, the Atomic Energy Commission and finally by the National Institutes of Health and the National Science Foundation. Such support was essential to obtain the personnel, equipment and supplies needed by the new fields of molecular and microbial genetics which flourished and indeed flowered during Beadle’s chairmanship.
During this third period there were many postdoctoral research fellows in the Biology Division, including S. Benzer (Crafoord Prize in 1993), who was a post-doctoral fellow in Delbrück’s group from 1949 to 1951, and was later recruited in 1967 as full professor. J. Weigle was a visiting professor and a valuable member of the Delbrück group. There were visits by F. Jacob (Nobel Laureate, 1965) and J. Watson (Nobel Laureate, 1962). B. McClintock returned in 1946 for a short visit, working with one of the graduate students, Jessie Singleton, perfecting a method of analyzing the chromosomes of Neurospora. Interestingly, R. Feynman, Caltech professor of physics (Nobel Laureate in Physics, 1965), spent part of an academic year working with R. Edgar and other members of the Delbrück group.

George Beadle at blackboard.
Courtesy of the Caltech Archives. © California Institute of Technology. All rights reserved. Commercial use or modification of this material is prohibited.
Beadle had remarkably versatile skills. He early abandoned his research on Neurospora in order to devote full time to being chairman. He was very successful in finding donors to endow postdoctoral fellowships and new buildings. The fellowships were often used to support visits by foreign scientists who otherwise would not have had been able to come to the USA. As in the previous period, teaching loads were kept light and much teaching was conducted in the form of seminars and journal clubs. The biology faculty was by and large a harmonious group and students were allowed considerable freedom to choose their professors. As one of a number of measures of the success of this atmosphere, the Nobel Prize in Physiology or Medicine was awarded to Professors Delbrück, Dulbecco and Sperry, as already noted, and in my case as well, for work carried out in the Division under the leadership of Beadle.

George Beadle and students.
Courtesy of the Caltech Archives. © California Institute of Technology. All rights reserved. Commercial use or modification of this material is prohibited.
Notes
1. Mutant and Wild Fly
 Photo kindly provided by Nils Ringertz.
Photo kindly provided by Nils Ringertz.
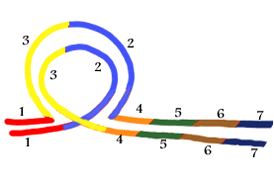
2. Inversion
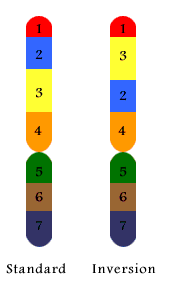 |
A chromosome having the standard sequence of regions labelled 1 to 7, inclusive, is shown on the left. If a segment labelled 2 and 3 becomes inverted then the new configuration shown on the right is called an inversion. When the standard and inverted chromosomes pair with one another the result is a chromosome loop as shown in the lower part of the diagram. |
 |
|
References
1. Allen, G., Thomas Hunt Morgan, pp. 1-447, Princeton University Press, Princeton, N.J. (1978).
2. Sturtevant, A.H., A History of Genetics, pp. 1-165, Harper and Rowe, New York (1965).
3. Goodstein, J.R., Millikan’s School, W. W. Norton and Co., New York. pp. 1-318 (1991).
4. Horowitz, N.H. Biographical Memoirs, vol. 59, pp. 26-52, National Academy Press, Washington, D. C. (1990).
First published 20 April 1998
Nobel Prizes and laureates
Six prizes were awarded for achievements that have conferred the greatest benefit to humankind. The 14 laureates' work and discoveries range from quantum tunnelling to promoting democratic rights.
See them all presented here.

